体軸形成(これまでの研究)
母性wnt8a転写産物のゼブラフィッシュ体軸形成における役割
ゼブラフィッシュの初期発生において、背腹軸の形成機構は受精直後から開始します。 卵黄植物極に存在する背側決定因子が、微小管依存性に将来背側へと輸送され、Wntカノニカル経路を介して背側特異的遺伝子の発現を誘導することで背側軸を決定すると考えられてきました。 wnt遺伝子の内、唯一wnt8aのmRNAだけが卵母細胞の植物極に局在し背側誘導能を持つことから、Wnt8aが背側決定因子の有力候補とされてきました。 本研究では、TALEN法によりwnt8a遺伝子変異体を作製しました。 ゼブラフィッシュwnt8a遺伝子は、染色体上並列にならぶ二つのORF(1/2)を有しています。 ORF1またはORF2単独の接合体変異体では体軸形成異常を示さないが、ORF1/2両方を欠損したwnt8a接合体変異体は強い前方背側化表現型を示すことから、ORF1とORF2が腹側、側方及び後側の組織形成に冗長的に機能することが示されました。 従来の予想とは異なり、wnt8a母性変異体は体軸形成に異常を示しませんでした。 一方、母性接合体複合変異体では、接合体変異体に比べてより重篤な前方背側化表現型を示しました。 これらは、母性wnt8aが初期背側決定には必須ではないが、接合体wnt8aと協調し腹側、側方及び後部の組織形成に関与することを示しています。 本研究ではさらに、Wnt6aを新たな背側決定因子の候補として見出しました。
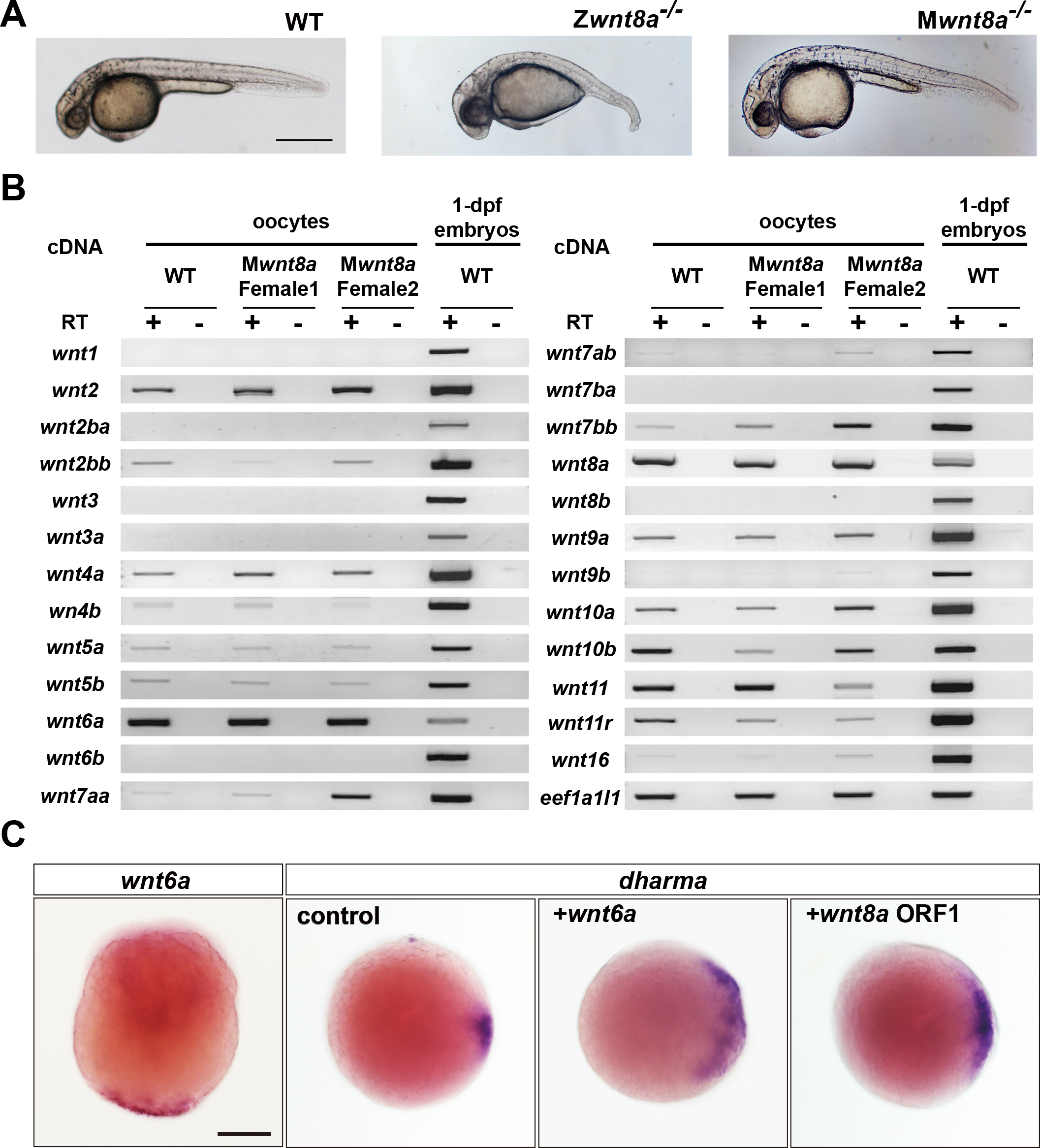
wnt8a欠損の表現型と新たな背側決定因子候補の探索。 (A)野生型(WT)およびwnt8a接合体(Z)、母性(M)変異体。1日胚。 (B)野生型(WT)およびwnt8a母性遺伝子欠損卵母細胞におけるwntファミリー遺伝子の発現(RT-PCR)。 1日胚はコントロール。 新たにwnt6aが母性に強く発現していることが示された。 (C)wnt6aの発現および機能解析。 wnt6aは卵黄植物極で発現しており、wnt8aと同様、背側特異的遺伝子dharmaの発現を誘導することが示された。
Dnai2は、メダカにおいて左右軸および腎臓形成に必要である
繊毛の機能欠損は、多発性嚢胞腎症や原発性繊毛機能不全症など様々な病気を引き起こすことが知られています。 私達の研究室で単離された左右軸形成異常を示すメダカ変異体miiを解析したところ、 その責任遺伝子が繊毛関連遺伝子のひとつであるdnai2aであることがわかりました。 この変異体では体の左右軸形成に必須なクッパー胞でのノード流が失われており、また、クッパー胞の繊毛では外腕ダイニン(ODA)が欠損していました。 一方、miiでは腎臓の繊毛の運動性およびODAに異常はなく、多発性嚢胞腎の表現型も検出されませんでした。 メダカでは種特異的な遺伝子重複により、dnai2aとは別にdnai2bが存在していました。 dnai2bはクッパー胞では発現していませんでしたが、腎臓ではその発現が観察されることから、腎臓ではDnai2aの欠損をDnai2bが補っていると考えられます。 実際、dnai2aとdnai2bの両方の機能を阻害することにより、多発性嚢胞腎の表現型が現れることがわかりました。 これらのことからDnai2はメダカにおいて繊毛の運動性、およびODA形成に必要であることが示されました。
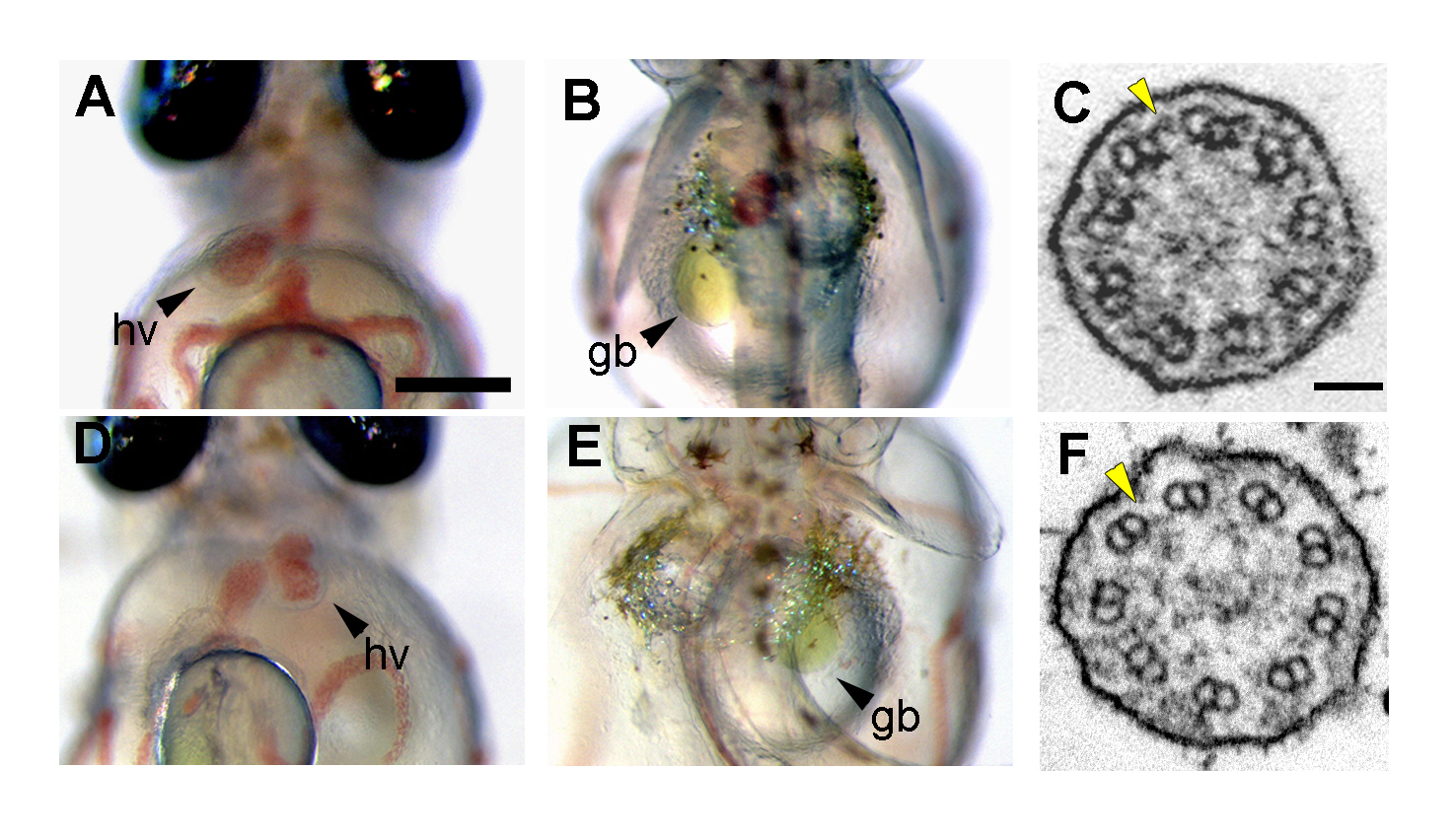
(A, B, D, E)受精後7日目の内臓の左右性。 (A, B)野生型胚では心室と胆のう(肝臓)は体の左側に位置している。 (D, E)mii変異体の中には内臓の逆位を示すものがいる。(A, D)腹側から観察。 (B, E)背側から観察。hv:心室、gb:胆のう。(C, F)クッパー胞の繊毛の電子顕微鏡像。 (C)野生型の繊毛は横断面図でみると外腕ダイニン(黄色の矢じり)を持った9+0構造をしていた。 (D)mii変異体の繊毛では外腕ダイニンが完全に欠損していた。