神経回路機能ー恐怖応答学習
ゼブラフィッシュ小脳において、顆粒細胞は古典的条件付け恐怖反応からの回復を制御している [Pubmed]
これまで小脳は古典的恐怖条件付けに関与することが示されてきたが、小脳のどの神経回路素子がどのように古典的恐怖条件付けに関わっているかは未だ明らかになっていません。私達はこの問題を解決するため、LED照明の消灯を条件刺激(CS)、電気ショックを無条件刺激(US)とした古典的恐怖条件付けをゼブラフィッシュ仔魚に対して行いました。USは拘束下のゼブラフィッシュ仔魚において、徐脈を引き起こしました。CSとUSを組み合わせて繰り返し提示すると、かなりの比率のゼブラフィッシュ仔魚で、CS提示に誘発された徐脈反応を示しました。私達は、古典的恐怖条件付け学習におけるゼブラフィッシュ小脳の役割を調べるため、小脳ニューロンにボツリヌス毒素やカルシウムインジケーターGCaMP7aを発現させました。ボツリヌス毒素により小脳体の顆粒細胞の神経伝達を阻害した場合、CS提示により誘発される徐脈反応は抑制されませんでしたが、むしろ徐脈反応は延長しまた。また、条件付け後にCS提示により活動する小脳ニューロンを特定しました。これらのニューロンの活動は、条件付けの間次第に上昇し、その後CSのみの提示を繰り返すと徐々に下降しました。これらのニューロンの多くはCS提示に対して即座に活動したが、いくつかのニューロンは遅れて活動しました。私達の発見により、少なくともゼブラフィッシュにおいて、顆粒細胞は恐怖条件付け反応からの回復を制御していることが示されました。
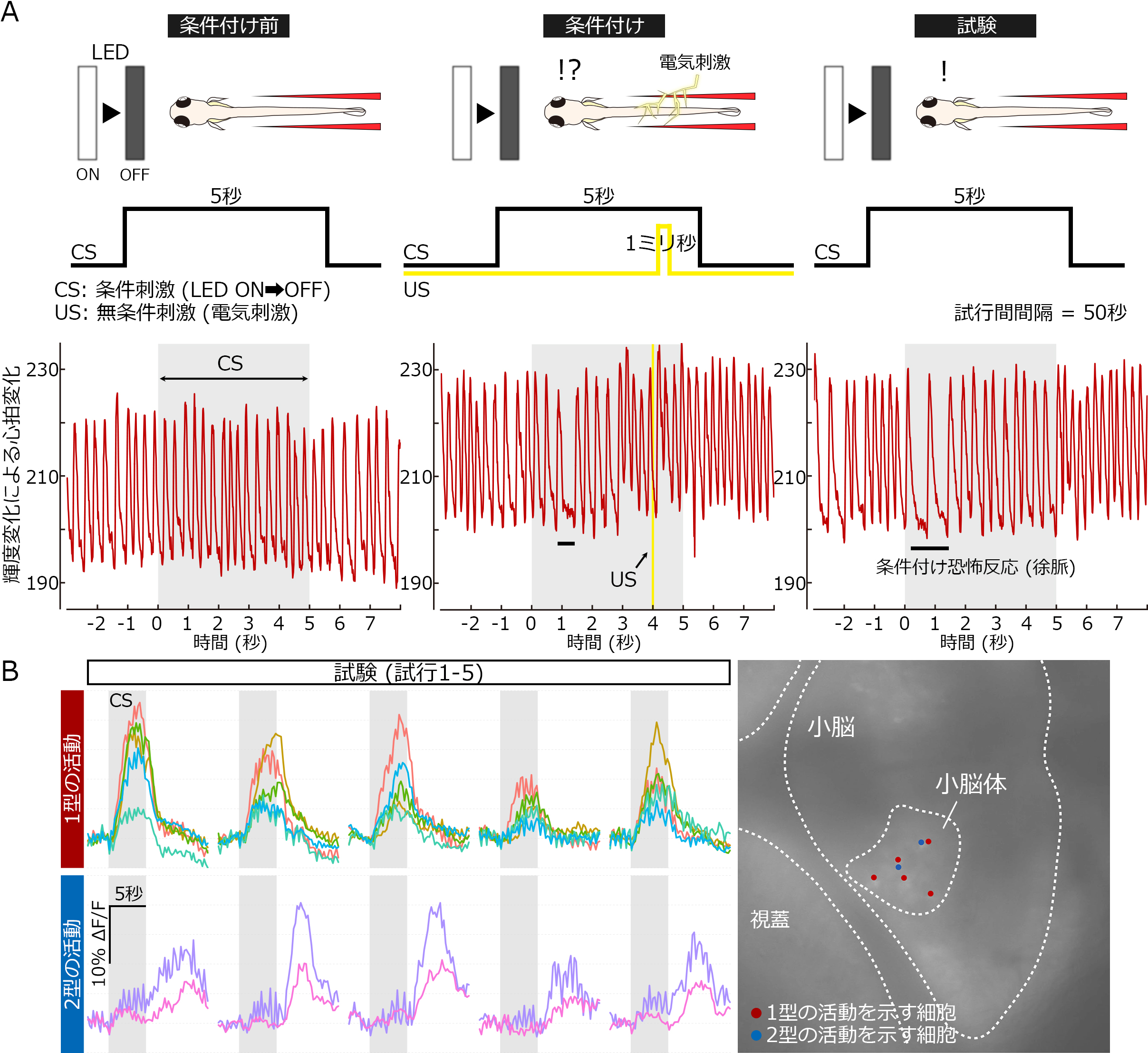
ゼブラフィッシュの恐怖応答学習。(A)恐怖応答学習のパラダイム。条件刺激(CS:LEDの消灯)と無条件刺激(US:電気ショック)を繰り返し与えると、CS誘発性の徐脈が観察される。(B)学習によって活性化される小脳ニューロン。GCaMP7a発現トランスジェニックフィッシュを用いた解析から、CS依存的な活動パターンには2種類あることが分かった。これら古典的恐怖応答学習において活動ニューロンは、小脳体と呼ばれる領域に存在し、顆粒細胞であることが示唆された。
メダカとゼブラフィッシュcontactin1b変異体は運動協調性を制御する神経回路の理解のための良いモデルを提供する [Pubmed]
メダカroは、遊泳中にふらつき、回転する運動異常を示す突然変異体です。 ポジショナルクローニングにより、ro遺伝子座は免疫グロブリンスーパーファミリータンパク質Contactin1b(Cntn1b)をコードすると予想されました。 そこで、CRISPR/Cas9法によりメダカcntn1b変異体を作製しました。 cntn1b変異体およびcntn1bとroのトランスヘテロ変異体はroと同様の遊泳異常を示したことから、roの責任遺伝子はcntn1bと同定されました。 また、TALEN法によりゼブラフィッシュcntn1ファミリー遺伝子変異体を作製しました。 cntn1a変異体では異常は見られませんが、cntn1b変異体において遊泳異常が観察されました。 このことから、Cntn1bは、真骨魚類において、遊泳運動に関して保存された役割をもつことが示されました。 Cntn1ノックアウトマウスは小脳神経回路に異常を示しますが、メダカ・ゼブラフィッシュcntn1b変異体においては小脳回路の形態異常は見出せませんでした。 しかし、メダカro変異体では、視運動性反応(OKR)の学習が低下し、水流に対する走性が不安定でした。 これらの結果は、Cntn1bが小脳神経回路を含む運動協調に関与する神経回路形成に関与することを示唆しており、 メダカ・ゼブラフィッシュcntn1b変異体は運動学習や運動制御について理解する良いモデルと考えられました。
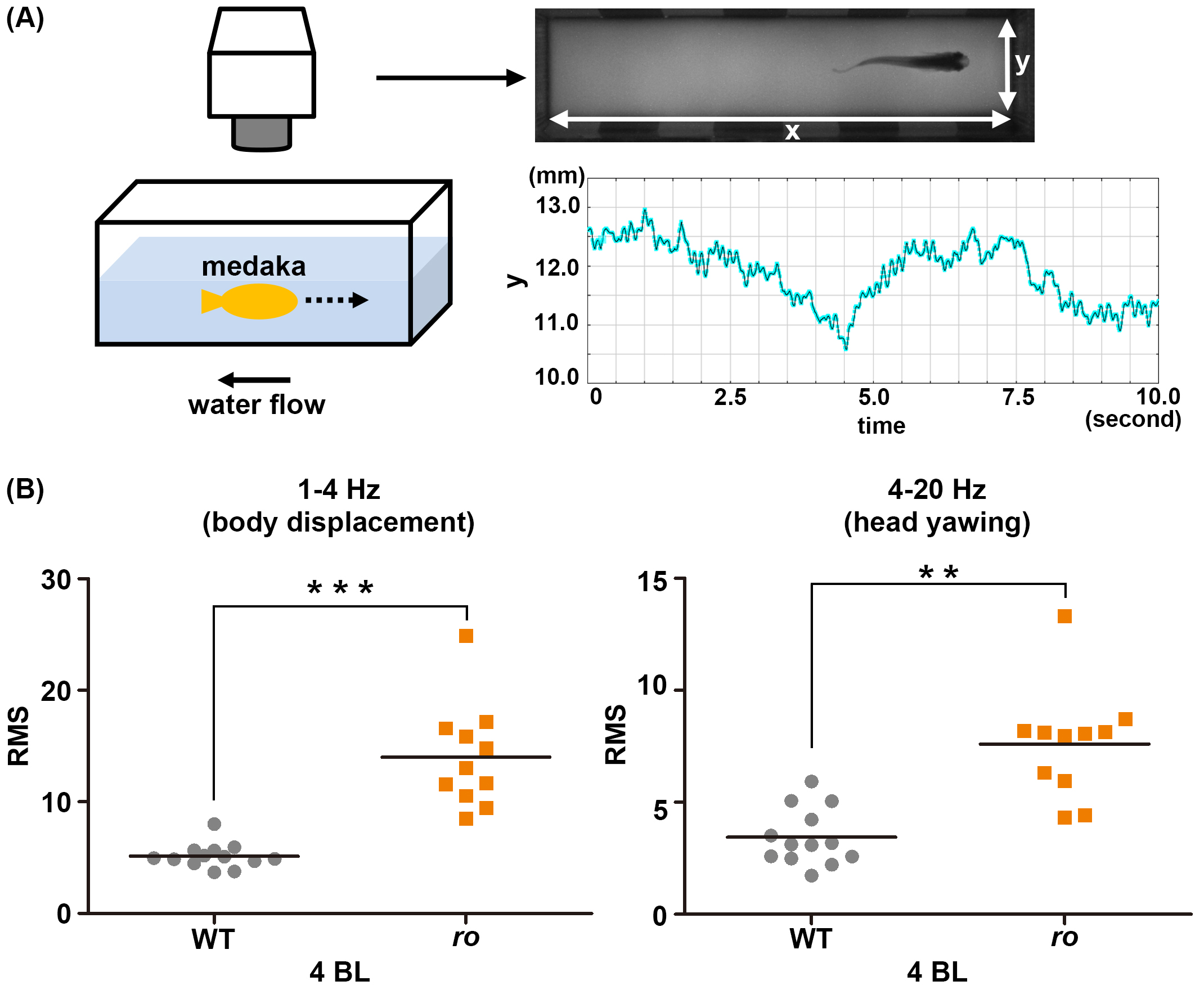
Cntn1bは走流性に必要である。(A) メダカは水流に逆らって泳ぐ性質をもつ。水流に対する遊泳行動をビデオ撮影し、頭部のy軸方向の動きをプロットして数値化し、x軸を時間としてグラフとして示した(右下)。(B) y軸の値について、Lab Chart softwareを用いて、low-frequency (1-4 Hz, body displacement: 体の移動を表す)とhigh- frequency (4-20 Hz, head yawing: 頭部のブレを表す)の動きとして分離し、二乗平均平方根(RMS)を計算した。 野生型(WT)に対して変異体(ro)では値が上昇した結果から、cntn1b変異体の遊泳が不安定であることが示された。