神経堤細胞分化ー色素細胞分化
Sox5とSox10はメダカとゼブラフィッシュの色素細胞の運命決定において異なる相互作用を示す [Pubmed]
私達は、メダカの色素細胞発生をモデルとして、SOXをはじめ種々の転写因子が多様な細胞種を作るメカニズムを研究してきました。 これまで、メダカの黄色素細胞と白色素細胞の系譜分岐においてSox5が運命選択を担う分子スイッチとして機能することを報告してきました。 本研究では、色素細胞発生への関与が示されていたSox10とSox5が相互作用する可能性を、メダカとゼブラフィッシュを材料として検証しました。
メダカは黒、虹、黄、白色の4種の色素細胞を持ち、ゼブラフィッシュは白以外の3種類を持っています。 Sox10は両魚種が共通に持つ3種類の色素細胞の発生に必須であり、Sox5はゼブラフィッシュではSox10の作用をわずかに阻害することが分かりました。 一方、メダカでSox5は黒色と虹色の発生においてSox10に阻害的でしたが、黄色と白色ではSox5とSox10の作用は協調的で、黄色の形成を促進し白色の形成を抑制していました。
本研究では、メダカにおいてSox5とSox10は系譜によって異なる作用を示すことが明らかになりました。 メダカの黄・白色素胞系譜に特有のSoxの機能は、ゼブラフィッシュと異なりメダカでは黄色の前駆細胞からさらに系譜を分岐させて白色を作ることと深く関係していると想像されます。 色素細胞発生に中心的な役割を果たすSox5とSox10の作用を少し変更することで、メダカは新奇な細胞種(白色)を進化的に獲得することができたのではないでしょうか。
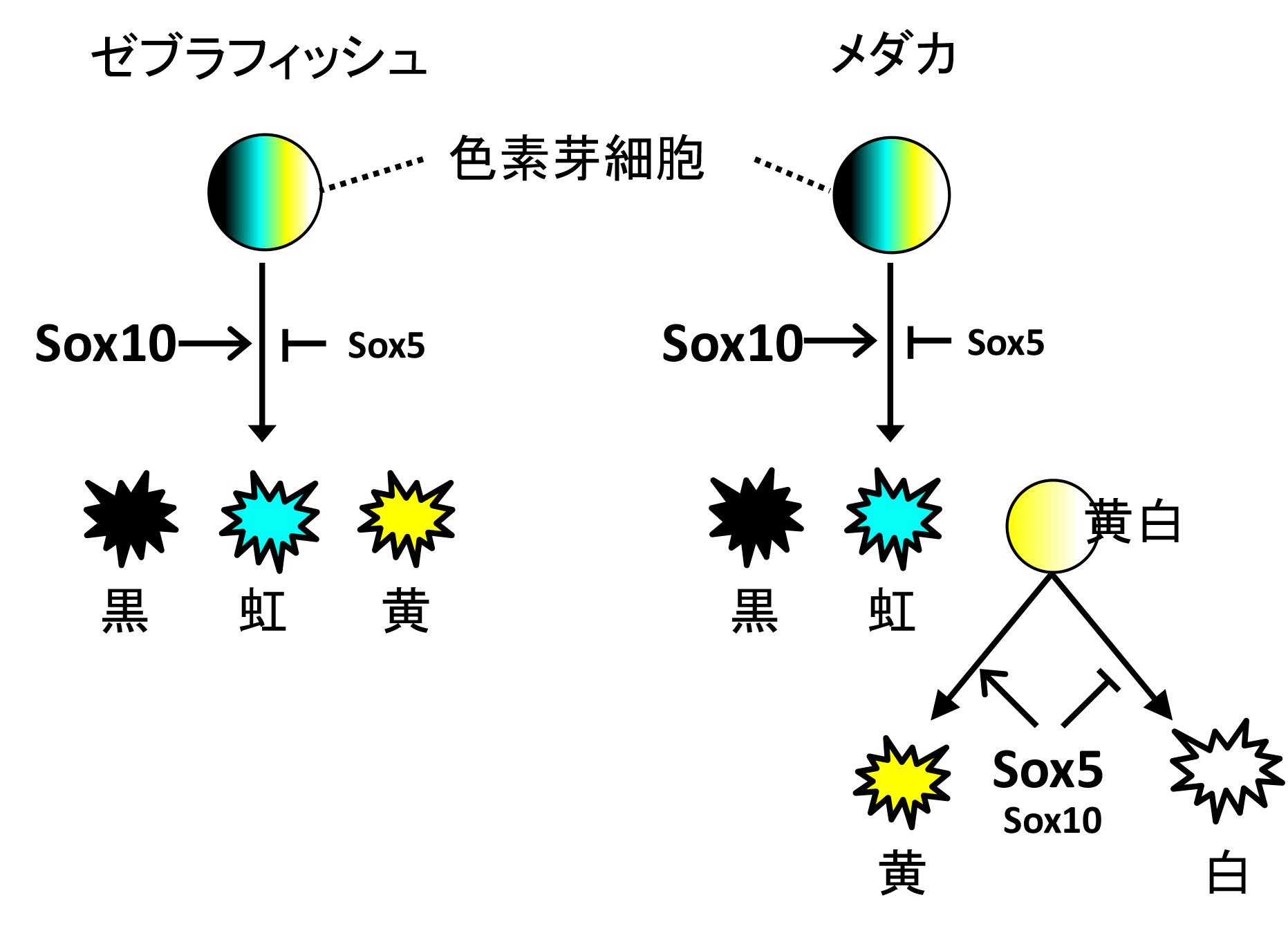
メダカとゼブラフィッシュの色素細胞系譜とSox5-Sox10の作用。 ゼブラフィッシュでは、黒、虹、黄色全ての分化に対してSox5はSox10の活性を阻害する。 メダカでは、Sox5は黒色と虹色の発生においてSox10に阻害的に働くが、黄色と白色ではSox5はSox10と協調的に働き、黄色の形成を促進し白色の形成を抑制する。
Sox5はメダカ色素細胞発生において運命決定スイッチとして機能する [Pubmed]
多能性幹細胞から多様な細胞が分化するメカニズムは正常発生に必要です。 神経堤細胞は、色素細胞を含む多様な細胞種を産生する多能性幹細胞です。 メダカは、神経堤細胞由来の四種類の色素細胞(黄色素胞、白色素胞、黒色素胞、虹色素胞)を持っており、多能性幹細胞から異なる細胞種が作られるメカニズムを理解する上で良いモデルを提供します。 メダカmany leucophores-3(ml-3)変異体は、黄色素胞欠損および過剰な白色素胞を有する変異体です。 私達は、ml-3の責任遺伝子が、神経堤細胞および分化中の黄色素胞に発現する転写因子Sox5をコードしていることを見出しました。 また、細胞移植実験から、Sox5は細胞自律的に黄色素胞分化を制御していることを明らかにしました。 転写因子をコードするpax7aは、神経堤細胞に発現しており、黄色素胞および白色素胞の両方の分化に必要であることが分かっています。 私達は、Sox5がPax7aの下流で機能すること示しました。 私達の研究成果から、多能性幹細胞である神経堤細胞から最初Pax7a依存性に黄色素胞と白色素胞の共通前駆細胞(中間前駆細胞)が作られ、その一部にSox5が発現しSox5依存性に黄色素胞を産生すると考えられました。本研究は、Sox5が黄色素胞と白色素胞の共通前駆細胞から黄色素胞分化を促進する運命決定スイッチとして機能することを示しています。
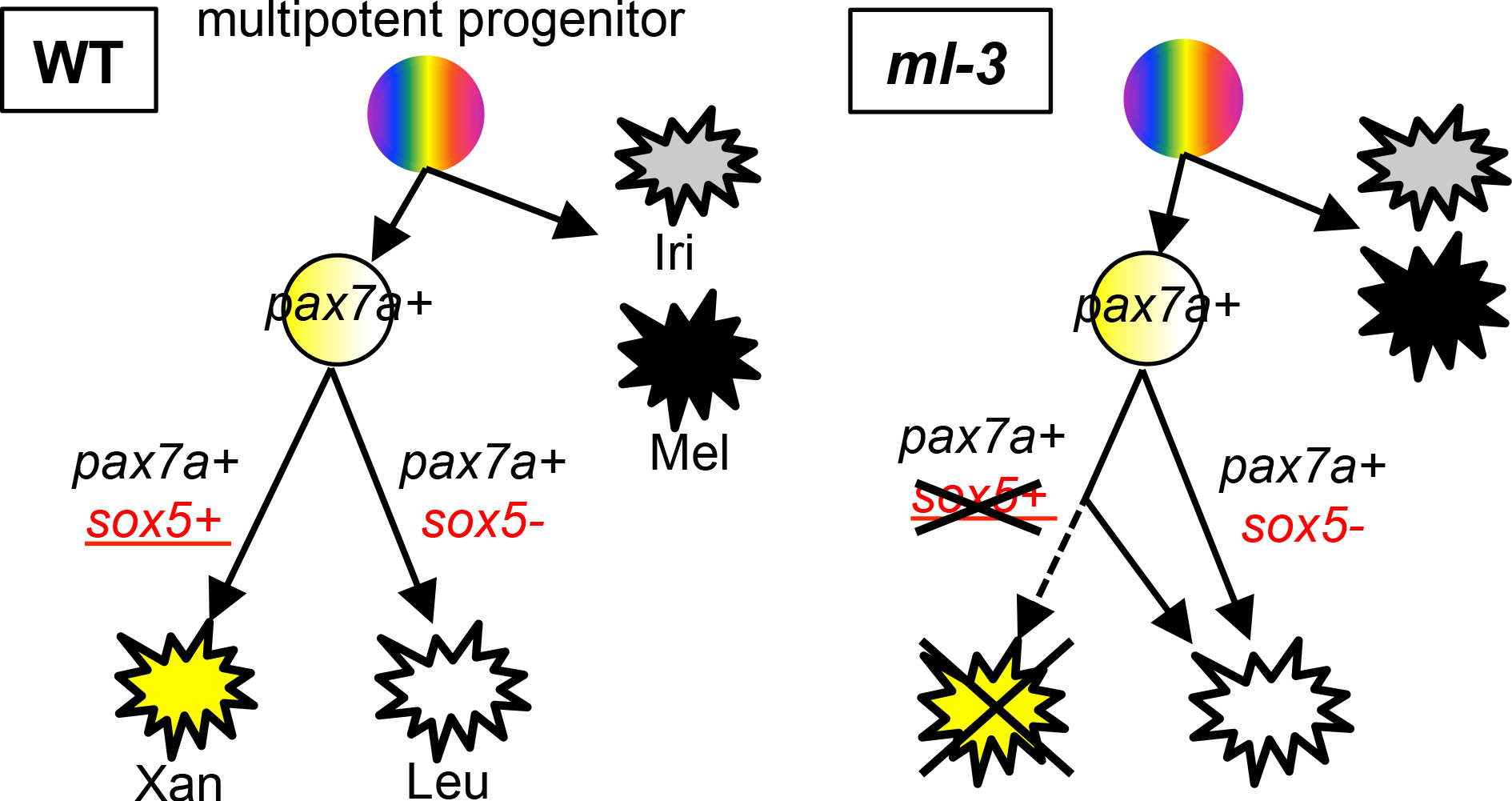
神経堤から黄色素胞(Xan)と白色素胞(Leu)への分化機構。黄色素胞と白色素胞は共通の前駆細胞から分化すると考えられる。 Sox5は白色素胞の代わりに黄色素胞に分化する運命決定に関わる。 共通前駆細胞の内、pax7a+ sox5+の細胞は黄色素胞に、pax7a+ sox5-の細胞が白色素胞に分化すると考えられる。 Sox5の欠損したml-3変異体では、黄色素胞への運命決定が起こらないため、全ての前駆細胞が白色素細胞へと分化する。 また、黒色素胞(Mel)と虹色素胞(Iri)の分化は、pax7aやsox5に依存しない。