
トピックス
体軸形成
これまでの研究
(1)背側オーガナイザー形成のメカニズム
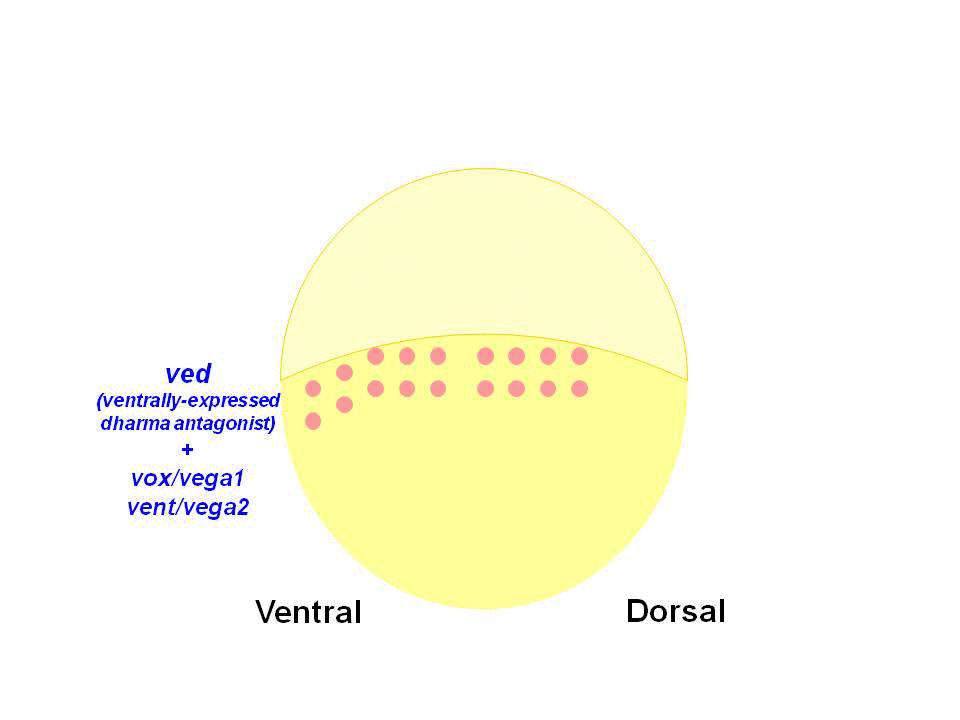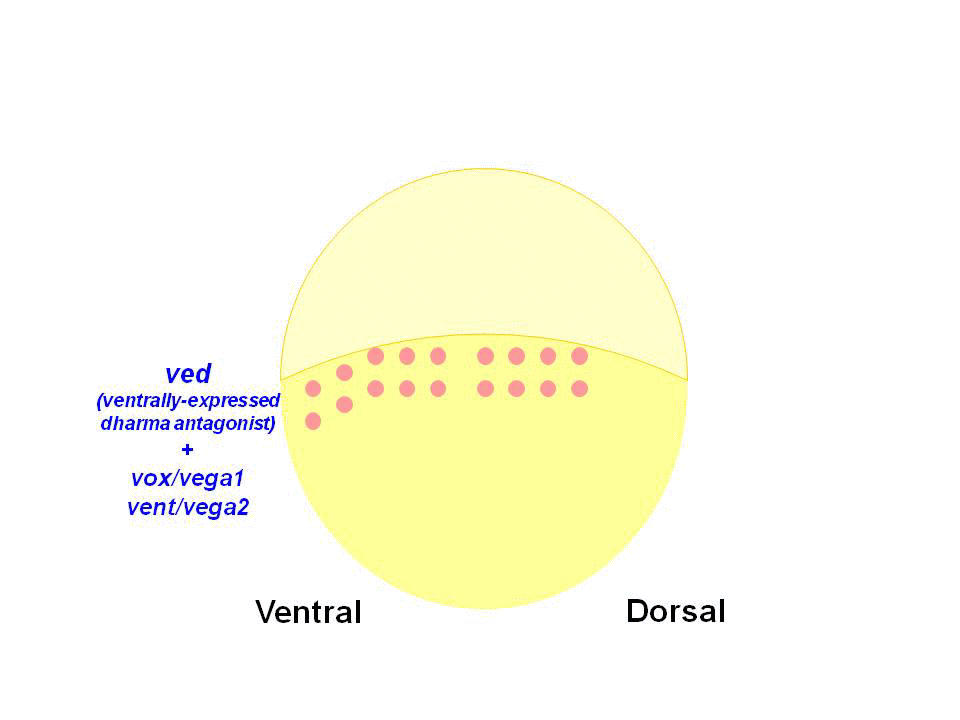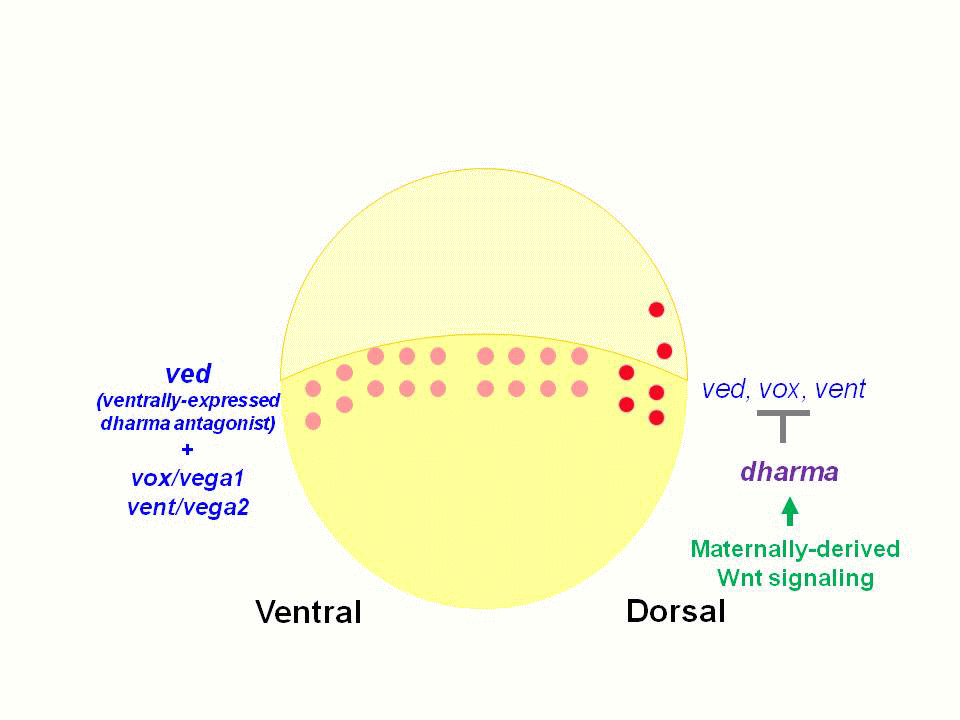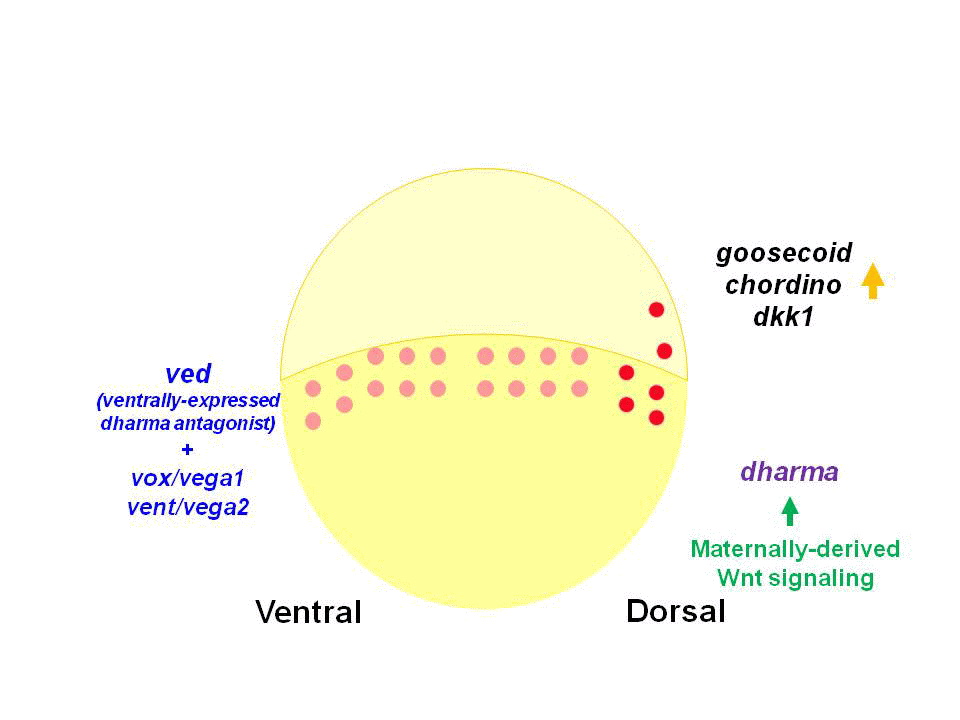
魚類・両生類では、受精卵植物極に存在する背側決定因子(Dorsal Determinants: DD)が胚背側に移動し、Wntシグナルを活性化すると考えられています。母性Wntシグナルは、背側胚盤細胞においてβ-cateninの核への蓄積を引き起こします。β-cateninは、転写因子Tcf/Lefと複合体を形成し、DNAの特異的配列に結合し、背側特異的遺伝子の発現を誘導します。
私達は、これまでWntシグナルの下流で制御されるホメオボックス遺伝子dharma (dha, bozozok変異体責任遺伝子)を単離し、dharmaが背側オーガナイザーの誘導に関与することを報告しました。またdharmaは、Wntシグナル下流で機能するnodal関連遺伝子ndr1(squint変異体責任遺伝子)と協調して、背側オーガナイザーを誘導することを見出してきました。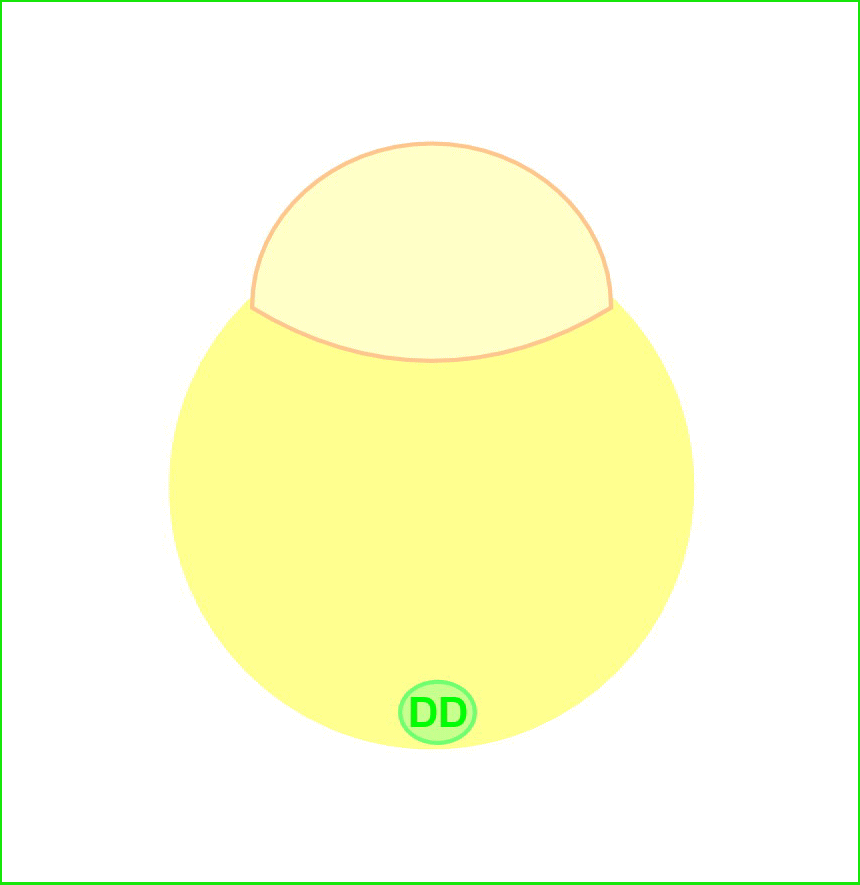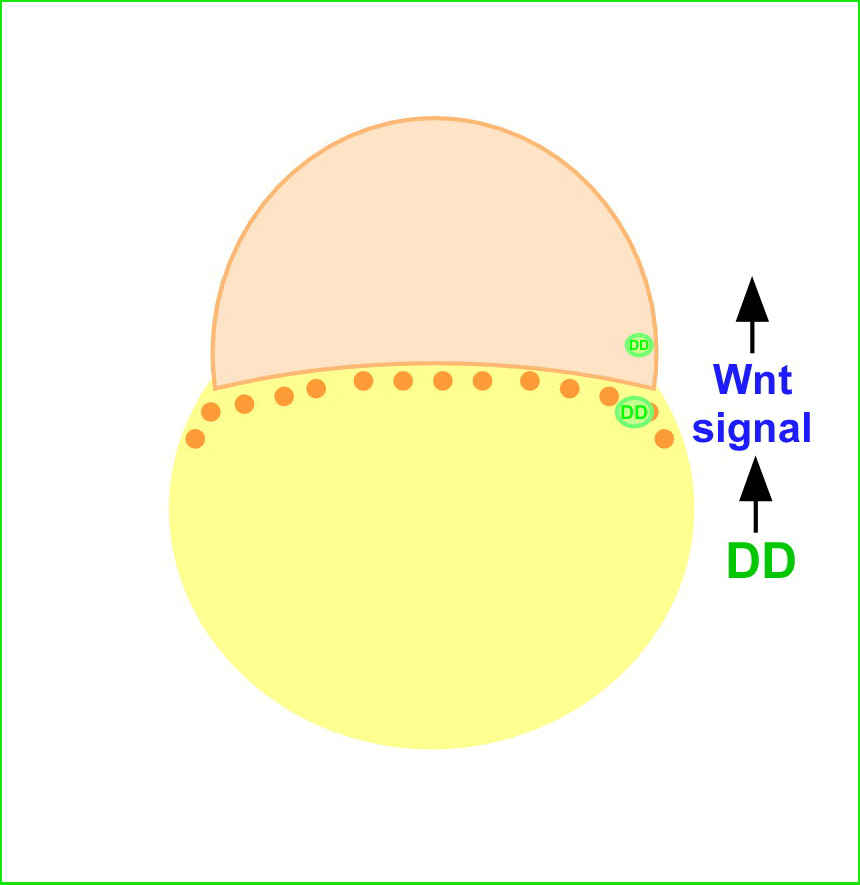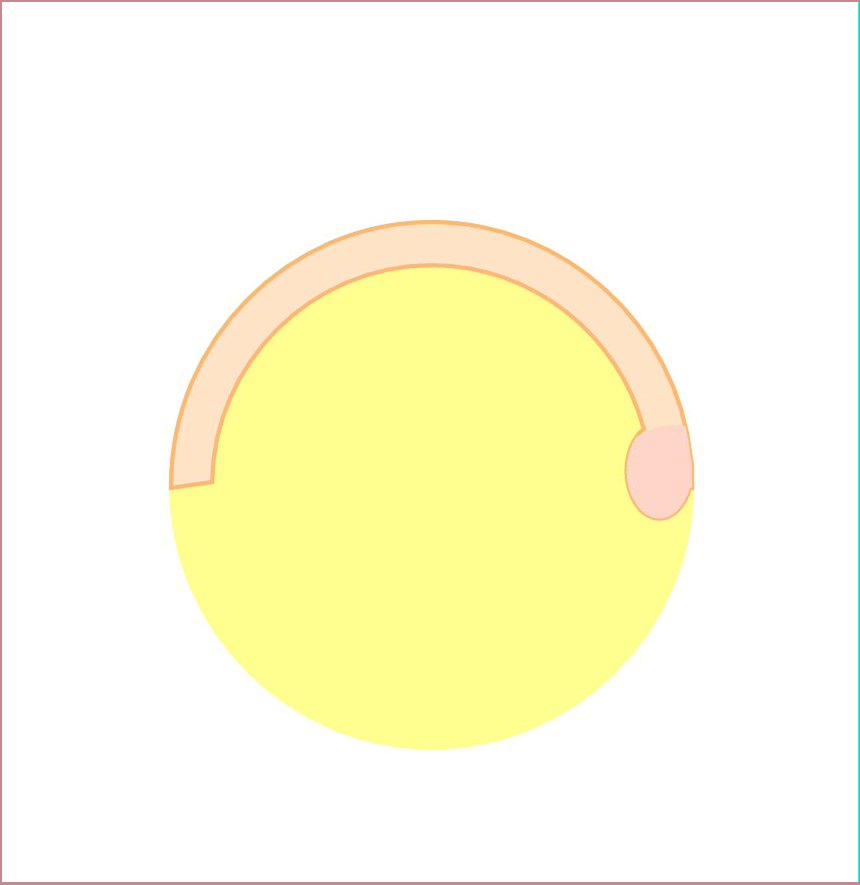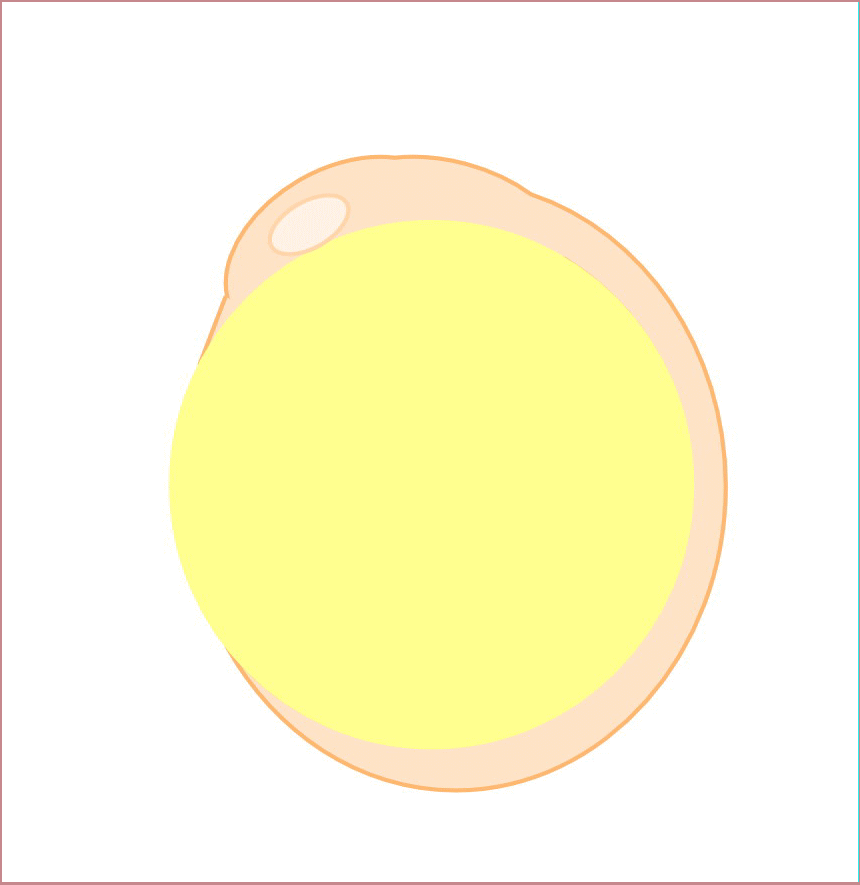
また私達は、Dharmaが転写抑制因子として機能し、腹側に発現するホメオボックス遺伝子vox, vent, vedの背側での発現を抑制していることを見出しました。Vox, Vent, Vedも転写抑制因子であり、腹側で背側オーガナイザーが形成しないよう機能しています。Dharmaは、この背側オーガナイザー形成の抑制を脱抑制することで、背側オーガナイザーを背側に限局させていると考えられました。
(2)オーガナイザーシグナルの制御
原腸胚においては、腹側に発現する腹側化因子Bmpと背側オーガナイザーから産生Bmp阻害因子(Chordin等)が相互作用することで、背腹軸形成が制御されていると考えられています。私達は、ゼブラフィッシュ腹側化変異体ogonの責任遺伝子を、Secreted Frizzled (Sizzled)と同定しました。SizzledはWnt阻害因子と類似の構造を持ちますが、Wntシグナルに作用するのではなく、オーガナイザー因子Chordinの分解酵素Bmp1を阻害することで、Chordinタンパク質はを安定化することを見出しました。Sizzledは、Bmpシグナル依存性に腹側に発現しBmpシグナルを阻害するフィードバック抑制因子です。Sizzledは、Chordinタンパク質の安定性を腹側で制御することにより、背腹軸形成に必要なBmpシグナルのグラディエント形成に関わっていると考えられます。
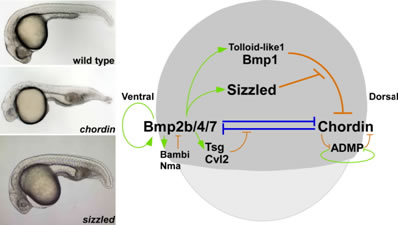
SizzledはTolloidファミリータンパク質分解酵素Bmp1/Tolloid-like1を阻害することにより、Bmp阻害因子Chordinの安定化を引き起こす。左は、野生型、chordin変異体、sizzled変異体(ogon)1日胚。chordin変異体は頭が小さく、腹側組織が拡大している。sizzled変異体においても、腹部組織拡大が認められる。
(3)左右軸形成
体の左右軸は、マウスの結節胞および魚のクッパー胞に存在する繊毛が回転し、流れを作り出すことが(ノード流)最初の引き金と考えられています。一方向のノード流は結果として、結節胞・クッパー胞領域左側でNodal関連分子の発現を上昇させ、それが左側側板中胚葉に伝えられ活性化されることで、臓器の左側化が起こると考えられています。私達は、クッパー胞の細胞で発現するNodal阻害因子Charonを同定しました。Charonはクッパー胞領域で右側が左側化しないよう機能していると考えられています。また、左右軸形成に異常を示すメダカ変異体miiを解析し、責任遺伝子dyenein axonemal intermediate chain 2a (dnai2a)が、左右軸形成に必要なクッパー胞の繊毛の運動に関与していることを明らかにした。
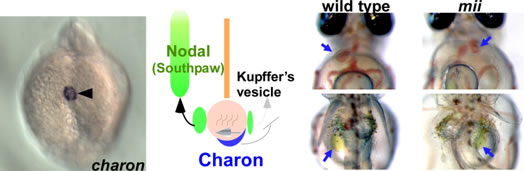
ゼブラフィッシュcharonの発現(左)とメダカmii変異体の表現型(右)。charonはクッパー胞の細胞に発現し、右側が左側化しないように機能している。Dnai2aを欠損しているメダカ変異体は、クッパー胞の繊毛が動かず、内臓逆位(心臓と胆のう)を示す個体が現れてきます。
現在の研究
私達は、背側形成に異常を示す母性遺伝子効果変異体tokkaebiを同定しました。tokkaebiホモ接合体雌魚から生まれた胚の多くは、全く背側組織が形成されません。tokkaebi胚では、Wntシグナルによるβ-cateninの蓄積が起こらず、またdharma、ndr1、背側オーガナイザー遺伝子の発現が減弱あるいは消失しています。tokkaebi変異体では、初期背側決定機構に異常があると考えられます。私達は、ポジショナルクローニングを行い、tokkaebi変異体の責任遺伝子が、微小管依存性のモータータンパク質Kinesinと貨物をつなぐリンカータンパク質Syntabulinであることを見出しました。私達の研究結果から、Syntabulinは微小管依存性の背側決定因子の輸送に重要や役割を果たしていると考えられます。Syntabulinが運ぶ背側決定因子の本態、および背側決定因子輸送の方向性を決める微小管形成のメカニズムを明らかにしたいと考えています。
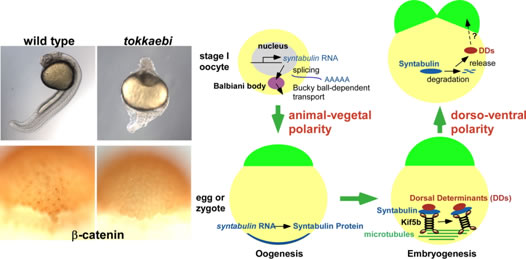
背側決定に関与するSytabulin。Syntabulin変異体tokkaebiは背側構造を持たず、また胞胚期の背側胚盤細胞におけるβ-cateninの核への蓄積が認められない(左)。Syntabulinを介した背側決定機構(右)。卵形成の過程においてsyntabulin RNAは、Balbiani bodyを通って植物極に移動し、タンパク質として翻訳される。受精後20分頃に形成された微小管上で、SyntabulinはKinesinと背側決定因子を結合させ、背側決定因子の運搬に関与しているのではないかと考えられる。