
トピックス
神経組織形
これまでの研究
(1)後方神経組織の形成
ゼブラフィッシュでは、原腸胚の胚盤周縁部にwnt3aとwnt8が発現します。Wnt3aとWnt8の機能を阻害した胚では、神経組織の前方化、体尾部の消失といった表現型が見られます。神経組織を含む体尾部の形成には、Wntシグナルの下流でcaudal関連遺伝子cdx1aとcdx4が機能することを見出しています。Cdx1aとCdx4の機能を阻害した胚では、脊髄の大半の組織は無くなりますが、驚いたことに神経組織の最後方に、前方に出来るはずの後脳の一部が形成されることが分かりました。このことは、Cdxは後方神経の運命決定を制御するだけでなく、前方神経が異所的に形成しないようブロックしていることを意味しています。
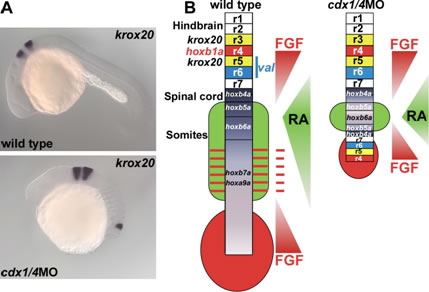
Cdx1a/4を欠損したゼブラフィッシュ胚では、後方神経が消失し、異所的に後脳が形成される。(A)cdx1a/4欠損胚では、菱脳節3、5番目に発現するkrox20が神経最後端にも異所的に発現する。(B)野生型およびcdx1a/4欠損胚でのFGFとレチノイン酸(RA)シグナルのグラディエントと神経パターンにング。cdx1a/4欠損胚の神経最後端では、FGFとRAシグナルがオーバーラップしており、Cdx非存在下ではこれらのシグナルにより後脳が形成されると考えられる。
(2)原始神経形成を制御する分子メカニズム
ゼブラフィッシュの後脳や脊髄では、発生初期に原始神経が形成されます。原始神経が形成される領域はプロニューロナルドメイン、その間を埋め原始神経が形成されない領域はインタープロニューラルドメインと呼ばれます。私達は、インタープロニューラルドメインに発現する遺伝子としてher3, her9を明らかにしました。her3, her9は、体軸位置情報をもとにプロニューロナルドメインを限局させる機能をもっており、プレパターン遺伝子として機能することを見出しました。また、原始神経形成に関与するホメオボックス遺伝子pnxを同定しました。
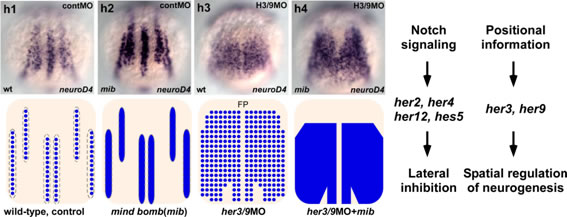
her3, her9はプレパターン遺伝子として機能する。野生型、Notchシグナル欠損胚mind bomb (mib)、her3/9欠損胚、およびNotchシグナルとher3/9両方が欠損した胚におけるneurod4の発現。Notchシグナル欠損胚では、プロニューロナルドメインの中で神経分化が亢進するがドメインの領域は大きな影響を受けない。一方、her3/9欠損胚では神経板全体がプロニューロナルドメインとなる。hes/her遺伝子の内、her2, her4, her12, hes5 (her15.1)はNotchのシグナルの下流で機能するが、her3, her9は後方神経においては位置情報に基づいてプロニューロナルドメインを決定しているものと考えられる。
(3)前脳形成に関わる遺伝子Fezf1, Fezf2
私達は、神経後方化に関わるWntシグナルを受けない、前脳および嗅神経に特異的に発現するジンクフィンガー遺伝子Fezf1 (Fez), Fezf2 (Fezl)を同定しました。ゼブラフィッシュのfezf2の変異体too fewは、間脳に存在するモノアミン(ドーパミン・セロトニン)作動性神経が減少しています。また私達は、Fezf1, Fezf2遺伝子のノックアウトマウスを作製し、哺乳類神経発生におけるFezf1, Fezf2の機能を解析しました。嗅神経に発現するFezf1が嗅神経の軸策伸長に、また嗅神経の標的である嗅球の形成に関与することを明らかにしました。また、Fezf2は大脳皮質第V層に位置する投射神経の運命決定に必須であることを見出しました。Fezf1, Fezf2は前脳で共発現しており、前脳の前後軸形成を制御していること、神経分化抑制因子Hes5の発現を抑制することで大脳皮質最初期の神経分化をコントロールしていることを明らかにしました。
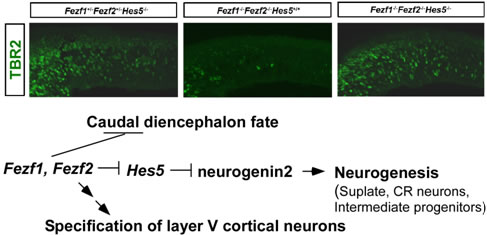
Fezf1, Fezf2の前脳神経発生における役割。Fezf1, Fezf2欠損マウスの前脳では、神経幹細胞からTBR2陽性中間神経前駆細胞への分化が抑制されているが、Fezf1, Fezf2, Hes5欠損マウスでは中間神経前駆細胞の形成が回復している(上段)。Fezf1とFezf2を介した前脳形成機構(下段)。Fezf1とFezf2は、Hes5を抑制することでneurogenin2を脱抑制し、神経分化を誘導する。Fezf1とFezf2は、前脳の前後軸形成を制御するとともに、大脳皮質5層の投射神経の運命決定に関与している。
現在の研究
私達は、ゼブラフィッシュとメダカを用いて小脳形成の研究を行っています。小脳は、後脳の前端背側に形成される神経組織で、円滑な運動を制御すると考えられています。最近の研究では、小脳は運動制御だけでなく認識・感情・学習などの高次な神経活動にも関与していることが分かっています。これまで、小脳の構造および発生はおもに哺乳類で解析されてきましたが、私達の最近の研究から哺乳類と魚の小脳の構造と発生は保存されていることが分かって来ました。哺乳類と魚の小脳において、プルキニエ細胞と顆粒細胞はそれぞれ主要なGABA作動性とグルタミン酸作動性神経であり、小脳外から入力線維を受けます。プルキニエ細胞は、脳室帯に存在するプロニューラル遺伝子ptf1aを発現する神経前駆細胞から発生し、下オリーブ核からの線維である登上線維を受けます。一方顆粒細胞は、菱脳唇に位置するプロニューラル遺伝子atoh1を発現する神経前駆細胞から発生し、苔状線維を受けます。苔状線維の信号は顆粒細胞からプルキニエ細胞に伝達されます。登上線維と苔状線維の二つの入力情報はプルキニエ細胞で統合され、小脳外へ出力されることで、高度な神経活動を制御していると考えられています。
私達は、プルキニエ細胞が分化する過程のライブイメージングを行い、プルキニエ細胞は最初多数の神経突起を持っているが、その内の一本が一次樹状突起として選択されることを見出しました。その過程で、ゴルジ装置の神経突起の根元への局在が、一次樹状突起の選択に重要な役割を演じている可能性を見出しました。さらに、細胞極性に関与するatypical PKCを介したシグナルが、ゴルジ装置の局在および一次樹状突起選択に必要であることを見出しました。
ゼブラフィッシュにおいては、受精後5日くらいまでに簡単な小脳神経回路を見ることができます。私達は、この点を利用して、小脳神経回路形成に異常を示すゼブラフィッシュ変異体のスクリーニングを行い、十以上の変異体を単離しました。また、名古屋大学でこれまで維持してきたメダカ変異体の中にも、小脳神経回路機能異常と思われる変異体がいます。このような変異体の責任遺伝子を同定し表現型を解析することで、小脳神経回路を形成する分子メカニズムに迫ることができればと考えています。
これらの研究を通して、ゼブラフィッシュ・メダカの小脳神経回路ニューロンに特異的に遺伝子を発現するシステムを構築しつつあります。近年開発されている光遺伝学などの技術を用いれば、ゼブラフィッシュとメダカの小脳がどのように情報を処理し、高次機能を発揮するかを明らかにできるのではないかと、夢を膨らませています。
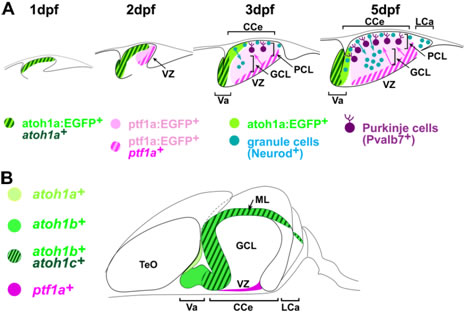
(A)ゼブラフィッシュにおける小脳ニューロンの発生。受精後1日目(dpf)に、atoh1a陽性細胞が上菱脳唇に形成。受精後2日目に、atoh1a陽性グルタミン酸作動性神経前駆細胞とptf1a陽性GABA作動性神経前駆細胞が、小脳原基の前半と後半部分を占めるようになる。受精後3-5日の間に、atoh1a陽性細胞は分子層(ML)でNeurod陽性の顆粒細胞に分化し、顆粒細胞層(GCL)に移動する。一方この間に、ptf1a陽性細胞は脳室帯(VZ)から背側に移動し、プルキニエ細胞に分化するとともにプルキニエ細胞層(PCL)を形成する。これらの過程を経て、受精後5日目には小脳三層構造が形成される。(B)ゼブラフィッシュの成体におけるプロニューラル遺伝子の発現。プロニューラル遺伝子の発現は、増殖神経前駆細胞で認められる。CCe:小脳体、LCa:小脳尾葉、TeO:視蓋、Va:小脳弁。
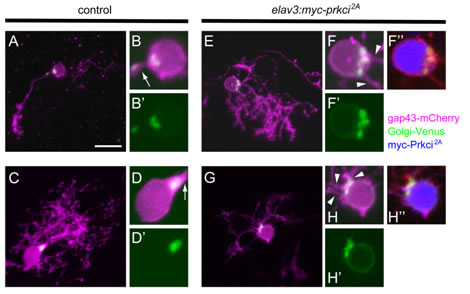
Prkciはプルキニエ細胞においてゴルジ装置の局在および一次樹状突起の選択に必要である。プルキニエ細胞特異的プロモーター(aldoca)により膜結合型mCherry(gap43-mCherry、紫)とゴルジ装置局在型Venus(Golgi-Venus緑)を発現させた受精後5日目のプルキニエ細胞。(E-G)はドミナントネガティブPrkci(Myc-Prkci2A、青)を発現させた細胞で、(A-D)はMyc-Prkci2Aを発現していないコントロールプルキニエ細胞。(B, B’, D, D’, F, F’, F’’, H, H’, H’’)細胞体近傍の強拡大像。コントロールのプルキニエ細胞では一本の一次樹状突起を伸長させており(矢印)、ゴルジ装置は一次樹状突起の根元に存在する。一方ドミナントネガティブPrkciを発現させたプルキニエ細胞は複数の一次樹状突起を持っており、ゴルジ装置の領域が拡大していた。また、増えた一次樹状突起は、拡大したゴルジ装置の近傍から伸長していた(矢じり)。