イネの側根の形が決まるしくみを時系列で明らかに
【本研究のポイント】
- ● イネの側根注1)の形(数、幅、長さ)が決まるしくみを、時系列で予測する回帰モデルを作成した。
- ● 側根の形態と、植物ホルモンのオーキシン注2)に関連する遺伝子の発現量の変化を時系列で比較して、側根の個性に関わる遺伝子の機能を解明した。
- ● 養水分の吸収を担う側根の個性を決めるメカニズムをより深く理解することで、高い養水分の吸収効率を持つ有用な品種の育成に貢献することが期待される。
【研究概要】
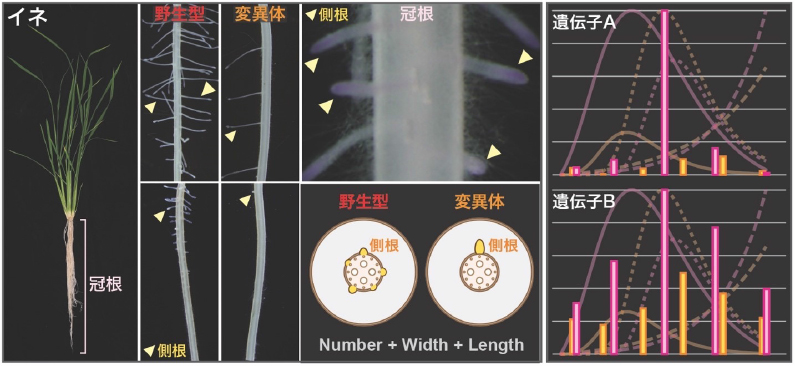
国立大学法人東海国立大学機構 名古屋大学生物機能開発利用センターの山内 卓樹 准教授は、同大学農学国際教育研究センターの犬飼 義明 教授、同大学大学院生命農学研究科の中園 幹生 教授らとの共同研究で、イネの冠根・不定根注3)から二次的に生じる側根の形態を3つの指標に分割し、将来側根を作る細胞が冠根で生じてからの時系列で解析することで、植物ホルモンのオーキシンを介した側根の個性の決定に重要なメカニズムの一端を明らかにしました。
現在、気候変動によるイネの収量・品質低下が課題となっており、収量の高い品種や乾燥耐性を持つ品種の作出が求められています。植物の側根は根茎の表面積を大きく増やすため、植物種間や栽培条件によって側根の形態の個性がことなる仕組みが分かれば、養水分の吸収を改良するような育種に役立つ可能性があります。
側根の数や大きさは植物ホルモンのオーキシンが制御していることが知られていますが、イネにおいてはオーキシン関連遺伝子がそのプロセスに果たす機能は完全には解明されていません。本研究では回帰モデルを用いて側根の形態を数、幅、長さに分けて時間経過でとらえることで、複数のオーキシン関連遺伝子が側根の形成に与える影響を明らかにしました。将来的には、本研究成果を活用し、側根の個性を決める遺伝子を制御することで、有用な作物品種の育成に貢献できる可能性があります。本研究成果は、2023年10月13日付アメリカ植物生物学会の学会誌「Plant Physiology」電子版に掲載されました。
【研究背景と内容】
側根は、作物の栽培に不可欠な土壌に直接触れる器官であり、根系注4)の表面積の増加に大きく貢献します。そのため、側根は圃場(ほじょう:作物を育てるための畑や水田)における養分吸収や乾燥土壌での水の効率的な吸収に重要な役割を果たします。側根のサイズは植物種間や栽培環境によって異なることが知られており、側根の質的な改良は作物育種における重要な観点の1つです。
側根原基注5)の形成やサイズ決定には植物ホルモンのオーキシンによるシグナル伝達注6)が関与することが知られていますが、作物のモデルであるイネの側根の個性を決める発生プロセスの理解を深めることで、側根の質的な改良が実現できます。
オーキシンシグナル伝達では、転写因子注7)であるAUXIN RESPONSE FACTOR(ARF)の機能を抑制する転写抑制因子注7)AUX/IAA(IAA)タンパク質がオーキシン依存的に分解されることでONになります(図1)。モデル植物のシロイヌナズナでは、主にARF7/19とIAA14によって側根や不定根の発生が制御されており、イネではARF19とIAA13が同様の機能を担うことが証明されつつあります。本研究では、IAA13の分解が阻害されることで、IAA13が発現する部位でオーキシンの有無に関わらずオーキシンシグナル伝達がOFFになるiaa13変異体を実験材料に用いました(図1)。iaa13変異体では、冠根から発生する側根の数が減少する一方で、側根の幅が大きくなることが知られていますが、その仕組みは完全には解明されていません。
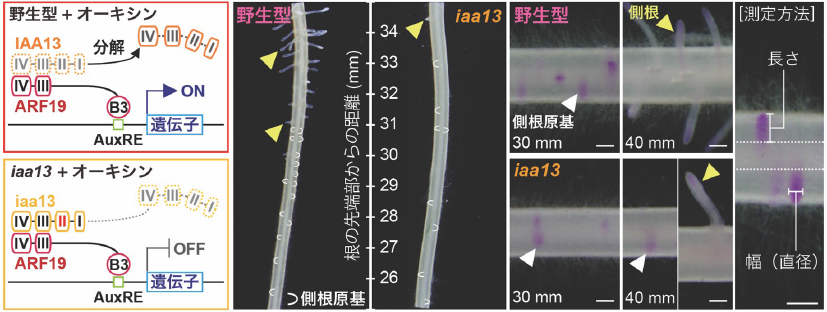
野生型では転写抑制因子IAA13がオーキシン依存的に分解されてARF19の機能がONになるが、iaa13変異体では分解されず、ARF19の機能がOFFの状態が維持される。それによって、オーキシンに依存した側根原基の形成や側根の成長が阻害される。一方、側根の幅は大きくなる傾向がみられた。ARFはDNA結合ドメイン(B3)でオーキシン応答配列(AuxRE)に結合して転写を制御する。
本研究では、側根の個性を決める遺伝子を探索するために、野生型のイネ(品種「台中65号」)とiaa13変異体の冠根の各部位における側根の数、幅および長さを計測しました。このとき、iaa13変異体の冠根は伸長速度が顕著に速いことから、野生型とiaa13変異体の冠根の先端部からの距離が同じであっても、将来側根を作る細胞が冠根で生じてからの時間(齢)は異なることが考えられました。この問題を解決するために、冠根の先端部からの距離を説明変数、各位置の側根の数、幅および長さを目的変数として回帰モデルを作成しました。側根の数および幅はゴンペルツ曲線注8)、長さについては指数曲線を用いました(図2)。続いて、冠根の伸長率(距離/時間)から、一定時間後の距離を算出し、その値を各モデルの関数の説明変数とすることで、時間(齢)に依存した側根の数、幅および長さを算出しました(図2)。さらに、時間毎の側根の数、幅および長さの増加率を可視化することで、iaa13変異体では側根の数と長さの増加が抑制されている一方、幅の増加率が長期間高い値を示すことを発見しました(図3)。
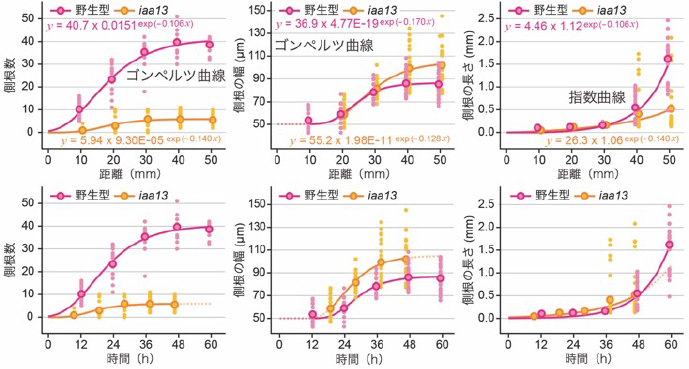
野生型とiaa13変異体の冠根の先端部からの距離を説明変数、各部位における側根の数、幅(直径)および長さ(図1参照)を目的変数とした回帰モデルを作成した(上段)。その上で、冠根の伸長率から各時間における先端部からの距離を算出し、各モデルの関数の説明変数として用いることで時間(齢)依存的なデータに変換した(下段)。小さい点が各サンプルの値、中心付近の大きな円が平均値を表す。iaa13変異体では、一部の側根は早期に伸長を開始するが、平均値としては野生型と比べて伸長の開始が遅れていた(下段右)。
続いて、オーキシン関連遺伝子の発現を回帰モデルを基に予測した時間毎の各側根の幅および長さの増加率と照らし合わせた結果、野生型ではARF11遺伝子の発現が高い値を示す一方、iaa13変異体では発現レベルが全体的に低下することが分かりました(図3)。実際に、ARF11の機能を失ったarf11変異体では、側根数の減少とともに、側根の伸長が抑制されました。このことから、シロイヌナズナのMONOPTEROS(MP)/ARF5とアミノ酸配列が類似するイネのARF11がMP/ARF5と同様に側根の発生や成長にも関与することが分かりました。
一方、オーキシンの細胞間の輸送に関わるPIN-FORMED(PIN)注9)タンパク質をリン酸化して、輸送方向を地上部側に変化させるPINOID(PID)注9)をコードする遺伝子の発現は、iaa13変異体で早い時期から長い時間高く維持されることが分かりました(図3)。この結果は、オーキシンが側根の先端部に供給されることが側根の伸長の開始に重要であり、iaa13変異体では側根の先端部へのオーキシンの供給が阻害されるため、側根の伸長の遅れと側根の幅の増加が起こることを示唆するものでした。
【本研究成果の意義】
今回の研究では、回帰モデルを利用して将来側根を作る細胞が冠根において生じてからの時間(齢)に依存した解析を行うことで、側根のアイデンティティの決定に転写因子やオーキシンの輸送方向を調節するタンパク質が関わっていることを明らかにしました。これらのタンパク質を適切に制御することで、側根の質的な改良による作物の養水分吸収能の強化を実現できる可能性があります。
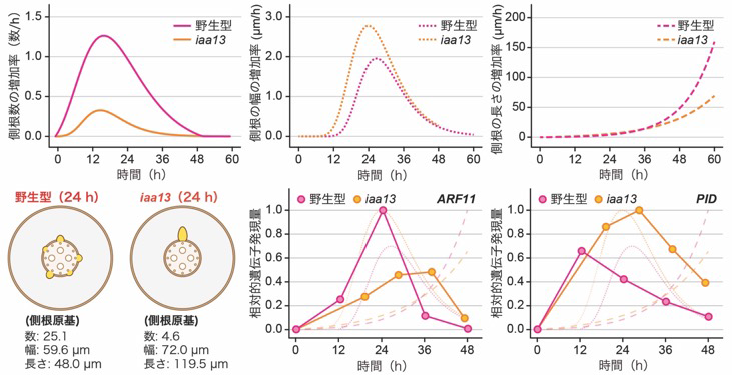
野生型とiaa13変異体の冠根における時間(齢)依存的な側根の数、幅(直径)および長さの増加率を比較した(上段)。野生型とiaa13変異体の24時間後の側根原基形成の模式図(下段左)。側根数は相対的な数で表現している。野生型とiaa13変異体の側根(原基)1つあたりのARF11とPIDの発現パターン(下段右)。
本研究は、文部科学省科学研究費助成事業学術変革領域(A)「不均一環境と植物」、新学術領域研究「植物の周期と変調」、科学技術振興機構 (JST) 戦略的創造研究推進事業(さきがけ)「フィールド植物制御」の支援のもとで行われました。
【用語説明】
注1)側根:
主根や不定根(冠根)、種子根などから枝分かれして側方に伸長する比較的細い根を側根と呼ぶ。側根の維管束系は発生した元の主根などと繋がっており、側根から吸収された養水分は主根などを経由して葉などの地上部に運ばれる。
注2)オーキシン:
植物において細胞の伸長や分化、分裂を促進することで、形態変化や、光・重力などへの応答を制御するホルモン。
注3)冠根(かんこん)・不定根:
幼根(胚発生時に形成される根で、双子葉類では後に成長して主根となる)や側根以外の根を主に不定根と呼ぶ。イネは節由来の複数の節根(不定根)から構成されるひげ根型根系を形成します。このイネの不定根を冠根と呼ぶ。
注4)根系:
植物の地下部を構成する器官全体の構造を根系と呼ぶ。イネ科植物の根系は、ひげ根型根系と呼ばれ、茎から生じる複数の節根(不定根または冠根)および節根から二次的に生じる側根によって構成される。
注5)側根原基:
主根などの中心部に存在する内鞘細胞が分裂を繰り返すことで形成される側根の基となる組織。イネ科植物では、内皮細胞が側根原基の一部として一時的に利用されることが知られている。
注6)オーキシンによるシグナル伝達:
転写(DNAをmRNAに写し取ること)のON/OFFを決めるタンパク質である転写因子ARFに結合して、その機能を阻害するタンパク質AUX/IAAがオーキシン依存的に分解されて調節されるシグナル伝達経路(図1参照)。
注7)転写因子・転写抑制因子:
ゲノムDNA上の特定の塩基配列を認識して結合し、近傍に存在する遺伝子の転写のON/OFFを制御するタンパク質を転写因子と呼び、特に転写を抑制する機能をもつものを転写抑制因子と呼ぶ。
注8)ゴンペルツ曲線:
代表的なシグモイド曲線の1つであり、個体や細胞の増殖を表す成長曲線として広く利用されている。1825年にBenjamin Gompertzによって、同一集団の死亡率の推移を示す関数として考案された。
注9)PIN-FORMED(PIN)・ PINOID(PID):
PIN-FORMED(PIN)タンパク質は細胞膜に局在して細胞外へのオーキシンの排出を担うキャリアータンパク質であり、オーキシンの輸送方向を決める重要な機能を担う。PINOID(PID)はPINの局在を根端方向から地上部方向に変化させるリン酸化酵素。
【論文情報】
雑誌名:Plant Physiology
論文タイトル:Age-dependent analysis dissects the stepwise control of auxin-mediated lateral root development in rice(齢依存的な解析によるオーキシンを介したイネの側根の段階的な発生プロセスの解明)
著者:Takaki Yamauchi, Akihiro Tanaka, Mikio Nakazono, Yoshiaki Inukai(山内 卓樹、田中 瑛大、中園 幹生、犬飼 義明)下線は本学教員
DOI: doi.org/10.1093/plphys/kiad548
URL: https://academic.oup.com/plphys/advance-article/doi/10.1093/plphys/kiad548/7311020?login=false