「緑の革命」遺伝子はジベレリンの合成酵素をコードしていた! |
私たちの研究テーマの1つは、イネの生長を制御する植物ホルモン、ジベレリン、に関するものだ。そのきっかけとなったのは、「緑の革命」に貢献した遺伝子を突き止めたことだった。この「緑の革命」の遺伝子については、有用農業形質保存研究室のホームページに詳しく書かれているので、そちらを参照して貰いたい。有用農業形質保存研究室との共同研究により、緑の革命に用いられた半矮性(背丈が低い)性質を持つsd1変異体は、ジベレリン合成に関与するGA20酸化酵素の遺伝子が機能しなくなった結果であることが解明された。
|
第二の「緑の革命」遺伝子を探せ! |
さて、活性型のジベレリンは6つの反応によって段階的に合成されて行く(図1)。GA20酸化酵素遺伝子はその5番目の反応を触媒する。sd1変異体が、ジベレリン合成不全によりイネの生長が阻害され矮性になったとしたら、それ以外の合成反応がダメになっても、同じようにイネの矮性が引き起こされるはずで、それらの矮性変異体を用いればsd1変異体を用いなくても、イネの収量を増大させることが期待できる。私たちはそう考えて、sd1以外のジベレリン合成変異体を調べ、残り5つについてそれぞれの機能と矮性形質の関係を系統的に解析した(図2)。その結果、残りの5つの反応についても、その反応がダメになるとイネは矮性を示すことが分かった。このことは矮性イネを作るために利用できる遺伝子はSD1遺伝子以外にもたくさんある(正確には残りの5種類)ことになるが、不思議なことに、イネの品種改良の歴史を見ると、実際の育種に利用された矮性遺伝子はSD1に集中している。唯一の例外は、かつて収量日本一に輝いた「短銀坊主」という品種のD35遺伝子だったが、d35変異体も現在は全く利用されていない。
|
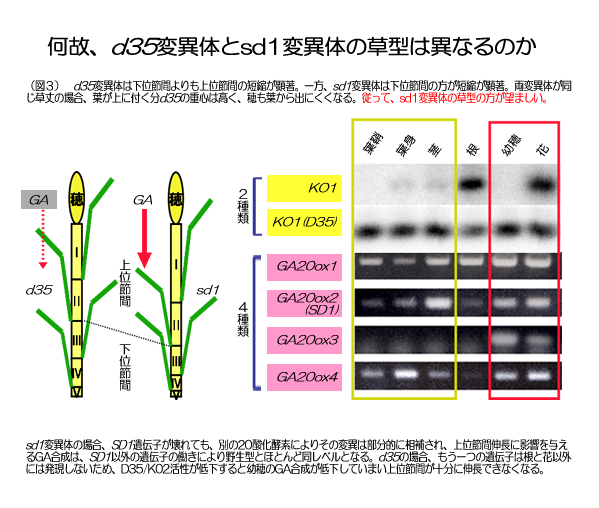
なぜd35変異体は利用されなくなったのか。日本で栽培されるイネの茎には5つの節間(Ⅰ~Ⅴ)があり、その上に穂ができる。sd1もd35も正常なイネより節間が短くなるため草丈が低くなるが、両者には節間の伸び具合に違いがある(図3)。sd1変異体は上位の節間に比べて下位が短い(下位節間の寸詰まり型)。逆にd35変異体は上位の節間が短くなる(上位節間の寸詰まり型)。この違いを育種の点から考えると、同じ矮性の草丈でも下位寸詰まり型のほうが有利だ。イネは各節から1枚の葉を出すので、下位寸詰まり型は重心が低くなり、より倒れにくい。さらに穂から出たすべての花が効率よく受粉するには、穂が葉に隠れない必要があるが、上位寸詰まり型では上の葉が穂を隠してしまい受粉がうまくいかない。
|
sd1もd35も同じジベレリン合成異常なのに何故このような違いを生じたのだろうか。これは、それぞれの遺伝子の発現パターンを考えると理解できる。sd1変異体の場合、SD1遺伝子が壊れても、イネに存在する別のGA20酸化酵素によりその変異は部分的に打ち消され、上位節間伸長に影響を与えるジベレリン含量は、SD1以外の遺伝子の働きにより野生型とほとんど同レベルとなる。d35の場合、もう一つの遺伝子は根と花以外には発現しないため、D35活性が低下すると穂のジベレリン量が低下してしまい上位節間が十分に伸長できなくなる。
|