ジベレリンはイネの生長を制御する重要な因子である! |
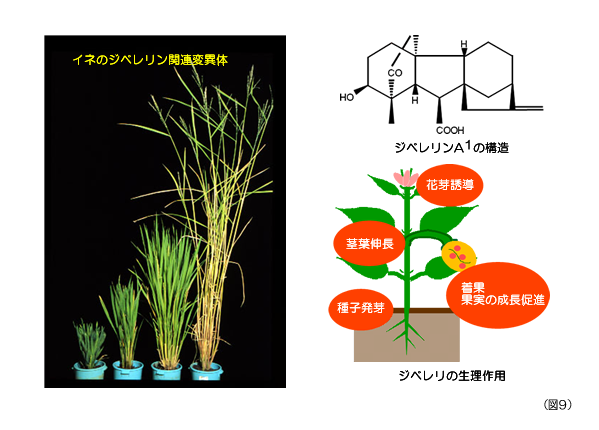
図9を見て欲しい。これは、植物成長ホルモンであるジベレリンの合成量やそれに対する感受性が変化したイネの変異体を集めた写真だ。イネはジベレリンというホルモンの合成や受容ができなくなると背丈が低くなり、逆にジベレリンを作りすぎたり、受容しすぎると背丈は高くなる。つまり、ジベレリンはイネの生長を制御する重要なホルモンなのだ。イネに多く含まれるジベレリンA1の構造を図9に示した。このジベレリンというホルモンは日本人の科学者によりイネの病気を研究する過程で見つけられた。現在、植物ホルモンと認められている化学物質は、ジベレリン以外に、オーキシン、サイトカイニン、アブシジン酸、エチレン、ブラシノステロイドの6種類があるが、日本人によって見つけられたホルモンはこのジベレリンだけだ。
ジベレリンの植物に対する作用は、図9右下に示したように、茎や葉の伸長を促進するだけでなく、それ以外にも色々な生理活性を示す。オオムギの発芽の際に胚から出たジベレリンが胚乳のアミラーゼの分泌を促し、発芽の際のエネルギーを供給することは、高校生物の教科書にも記載されており、特に有名だ。
|
イネの突然変異体を用いてジベレリンの研究をする! |
私たちはイネを研究材料に用いてジベレリンの研究を行っている。ジベレリンを合成できなくなったイネは背丈が低くなる。私たちは背丈が低くなった(矮化した)イネの突然変異体をたくさん集めて、その中からジベレリンに関係した変異体を選抜した。この方法で、ジベレリン合成に関わる6つの過程すべての変異体を見つけることに成功した。この6つの過程とそれぞれの変異体の写真は「半矮性化」の項の図1と図2に示した。私たちはこのジベレリン合成変異体収集の過程で、奇妙な変異体があることに気づいた。それは、形態はジベレリン合成変異体と非常に似ているにもかかわらずジベレリン合成変異体ではない、というものである。私たちがジベレリン合成変異体か否かを判断する場合、それがジベレリン欠乏により矮性の形態を示すことと、その矮性形態がジベレリンを人為的に添加することにより回復することを基準としている(図10左)。ところが、この奇妙な変異体は典型的なジベレリン欠乏の形状を示すのに、ジベレリンを添加しても、その矮性形質が回復しなかった(図10右)。私たちはこのような変異体をジベレリン不感受性矮性変異体(gibberellin insensitive dwarf, gid1変異体)と名付けた。
|
|
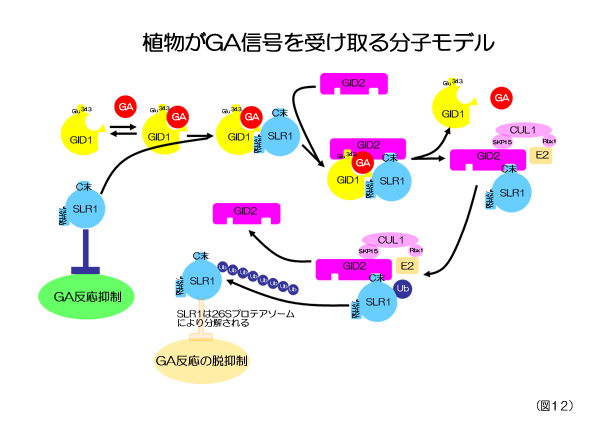
私たちはこれまでにジベレリン不感受性矮性変異体を2種類(gid1とgid2)見つけて、これらの変異体について実験を行ってきた(図11)。その結果、この2つの変異体はともに、ジベレリンの受容に関与する重要なタンパク質をコードしていることが分かった。すなわち、GID1はジベレリンの受容体を、GID2はジベレリン信号伝達を抑制する因子(この因子をDELLAタンパク質と呼ぶ)の分解に関わるタンパク質の情報を担っていることが分かった。そして、これらの変異体を用いた研究からジベレリンの受容メカニズムが明らかにされてきた。それをまとめたものが図12である。
|
GID1は核の中にあるジベレリン受容体である。細胞内にジベレリンが存在しない時、GID1は単独で存在している。このとき、ジベレリン反応はDELLAタンパク質(イネのDELLAタンパク質を特にSLR1と呼ぶ)により抑制されていて、その為、イネの生長は押さえられた状態にある。ここで、ジベレリン(図中ではGAで示されている)がやってくると、GID1とジベレリンが出会い、結合する。このGID1-GA複合体は、ジベレリン反応を抑制していたSLR1と結合することが可能となり、GID1-GA-SLR1という3つの分子からなる複合体を構成することになる。つまり、ジベレリンはジベレリン反応を止めていたSLR1と受容体GID1との結合を助ける役目をしていると言える。さて、GID1と結合したSLR1は、おそらくその立体的な構造を変えることにより、もう一つ別のGID2というタンパク質と結合出来るようになる。その結果、GID1-GA-SLR1-GID2という4つのタンパク質からなる複合体ができることになるが、実際には、このような複合体は観察されず、SLR1とGID2からなる複合体ができる。この理由はまだ分からないが、おそらくGID1-GA-SLR1-GID2という複合体は非常に不安定で、GID2が結合した瞬間に、GID1とジベレリン分子はこの複合体から離れていってしまうのだろう。実は、このGID2はタンパク質の殺し屋とでもいうべき働きを持っており、GID2が結合したタンパク質は、ユビキチンと呼ばれる短いペプチド分子を付加され、これを目印にタンパク質分解工場である26Sプロテアソームという所に運ばれ分解されてしまう。このGID2がSLR1にユビキチンを付ける反応には、SKP2、CUL1、Rbx1、E2等GID2以外に多くのタンパク質の関与が必要と考えられている。
このジベレリンの受容反応をまとめると、ジベレリン反応を止めているSLR1(DELLAタンパク質)が、ジベレリン受容体GID1と分解因子GID2の働きによりジベレリン依存的に分解され、その結果、ジベレリン反応が始まる、と言うことができる。このジベレリン反応のモデルは非常に興味深いことを示唆している。それは、このモデルは、イネ(もっと広げては植物はと言っても良い)はいつでも生長する状態にあり、その生長しようとする反応を無理矢理止めているのがDELLAタンパク質(イネの場合SLR1)で、実際に生長する場合は、この成長を止めているタンパク質を分解して、本来の状態に戻すことにより生長を促している、ということを意味しているからだ。
ここまで理解出来た人は、それではもし、イネにおける生長のブレーキ、つまりSLR1が壊れてしまったら、そのイネはどうなってしまうのか、と言う疑問を持ったかもしれない。実はこのようなイネの変異体が実際に存在する。イネの生長ブレーキ/SLR1が壊れると、予想通り、そのイネはひたすら生長するために異常にひょろ長くなり、すぐに倒れて死んでしまう(図13左下のイネ)。一方で、生長ブレーキを壊せなくなったイネがGID1やGID2が壊れたイネに対応し、イネは生長出来ないために強い矮性を示す(図13真ん中のイネ)。この場合、ブレーキを壊すことができないために、この変異体にいくらジベレリンを掛けても生長は始まらない、つまりジベレリン不感受性の矮性と言うことになる。これらの変異体が教えてくれることは、自動車の安全な走行もイネの正常な生長も同じで、止まりたい時はブレーキが掛かり、走りたい時にはブレーキが外せる、と言うことが非常に大切と言うことだ。 このような研究は、別のところで述べられている「すぐに農業に役立つ」研究ではない。しかし、私たちはこのような基礎研究も非常に重要だと考えている。というのは、基礎研究で見つかる新しい事実(上の場合はジベレリンの受容から、それに引き継ぐイネの生長メカニズム)から、これまで考えつかなかったような新しい技術が生まれる可能性があるからだ。実際、現在私たちは、この基礎研究で見つけたイネの生長ブレーキタンパク質、SLR1をうまく育種に利用できないかを考えており、その研究を始めた。この研究がうまくいくと、イネの「緑の革命」で用いられたジベレリン量を減少させることによりイネの草丈を短くして収量増加を達成する、という考えとは全く異なった戦略によりイネの草丈制御が可能となる。やはり、基礎研究から得られる基礎的な知識というのは、新しい技術革新には必要不可欠なのだ。
|